
Epífitos Vasculares sobre Diez Forófitos de las Yungas del Noroeste Argentino, con Énfasis en Helechos
Vascular Epiphytic on Ten Phorophytes from the Yungas of Northwestern Argentina, with a
Focus on Ferns
 Dalma G. Jaimez*1, Priscila D. López 2 & Adrián M. Jarsún1
Dalma G. Jaimez*1, Priscila D. López 2 & Adrián M. Jarsún1
1. Facultad de Ciencias Naturales, Universidad Nacional de Salta, Av. Bolivia
5150, Salta, Argentina. *jaimezdal@gmail. com; adrianjarsun@gmail.com
2.
Smithsonian Tropical Research Institute, Panamá. damarispl067@gmail.com
Resumen
Las epífitas vasculares son componentes clave en la riqueza de especies de los bosques
tropicales húmedos, donde contribuyen significativamente a la biodiversidad y a la estructura del ecosistema. El objetivo de este estudio fue evaluar la diversidad
de epífitos vasculares asociadas a diez especies de forófitos en la región biogeográfica de las Yungas, en el noroeste
de Argentina. Se muestrearon entre cinco y diez individuos por especie de forófito, con un diámetro
a la altura del pecho (DAP) superior a 30 cm, en cinco localidades: La Caldera (camino de
cornisa), Parque Nacional Calilegua, Potrero de Yala, Quebrada de San Lorenzo y Tiraxi. En total, se registraron 47 especies de epífitas, de las cuales 18 correspondieron a helechos. La familia Polypodiaceae presentó la mayor diversidad de helechos
epífitos. Las especies más
frecuentes en los forófitos muestreados
fueron Campyloneurum aglaolepis, Microgramma squamulosa y Pleopeltis
tweedieana. Los forófitos que albergaron la
mayor diversidad epifítica fueron
Cedrela angustifolia, Tipuana tipu, Erythrina falcata y Ocotea porphyria, mientras
que Nectandra angustifolia y Zanthoxylum coco presentaron la menor diversidad. Además, se observó
que el 76 % de las epífitas
habitan predominantemente en la copa interna de los forófitos.
Palabras claves: Bosque Nublado;
Forófitos; Polypodiaceae; Yungas.
Abstract
Vascular epiphytes are key components of the species
richness in tropical
rainforests, significantly contributing to both biodiversity and ecosystem structure. The aim of this study
was to assess the diversity of vascular epiphytes associated with ten phorophyte species in the Yungas biogeographic region of northwestern Argentina. Five to ten phorophytes with a diameter at breast height (DBH) greater than 30 cm were sampled
across five localities: La Caldera (camino de cornisa),
Calilegua National Park, Potrero de Yala, Quebrada de San Lorenzo,
and Tiraxi. A total of 47 epiphytic species
were recorded, 18 of which were ferns. The Polypodiaceae family exhibited the
highest diversity within the epiphytic ferns.
The most frequent species
found on the sampled
phorophytes were Campyloneurum aglaolepis, Microgramma squamulosa, and Pleopeltis tweedieana. The phorophytes
hosting the greatest epiphyte diversity were Cedrela angustifolia, Tipuana tipu, Erythrina falcata, and Ocotea porphyria, whereas Nectandra angustifolia and Zanthoxylum coco supported the lowest diversity. Additionally, 76% of the recorded
epiphytes predominantly inhabit the inner canopy of the phorophytes.
Keywords: Cloud Forest; Forophytes;
Polypodiaceae; Yungas.
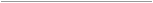
Jaimez, D. G., López, P. D., & Jarsun, A. M. (2025) Epífi Vasculares sobre Diez Forófi de las Yungas del Noroeste
Argentino.
Revista Ciencias Naturales, 3(1), 38-49. https://id.caicyt.gov.ar/ark:/s29535441/ouar2ujxm
Recibido: 6/2/2025 Aceptado: 24/6/2025 Publicado: 30/6/2025 Editor: Guillermo Terán
 Artículo de acceso abierto
bajo Licencia Creative Commons Atribución-No Comercial-Compartir Igual
4.0 Internacional (CC BY-NC-SA
4.0)
Artículo de acceso abierto
bajo Licencia Creative Commons Atribución-No Comercial-Compartir Igual
4.0 Internacional (CC BY-NC-SA
4.0)
Los epífitos vasculares desarrollan su ciclo de vida sobre otra planta, sin extraer agua o
nutrientes de ellas. Estas plantas dependen
de la humedad atmosférica, de la materia
orgánica que se acumula
en la copa de los árboles, así como de sus adaptaciones para captar y almacenar agua (Benzing, 1990; Wolf, 1993; Zotz, 2016).
A nivel mundial el epifitismo alcanza
el 9
% de la diversidad de plantas vasculares, tanto en el Neotrópico como en el Paleotrópico. Las epífitas vasculares,
como orquídeas, bromelias, aráceas y helechos, entre otras, son componentes clave
de la riqueza de especies de los bosques tropicales húmedos, donde contribuyen
significativamente a la biodiversidad y la estructura del ecosistema (Gentry & Dodson, 1987; Ibisch et al., 1996; Kersten, 2010; Bonnet et al., 2011; Zotz, 2013)
El éxito del
establecimiento para las
epífitas está limitado por las características de los árboles hospedantes o forófitos (Wagner et
al., 2015), como el tamaño de los árboles, forma y patrones de ramificación, composición química y morfología de la corteza. También
está relacionada con las condiciones de humedad del sustrato, patrones
fenológicos, estabilidad de la corteza y estructura del dosel
(Benzing, 1990; Hirata et al.,
2008; Laube & Zotz, 2006; Wagner
et al., 2015).
En los bosques húmedos tropicales, las epífitas son un componente ecológico
impor- tante, ya que contribuyen a la regulación hídrica, al ciclo de los nutrientes, a la riqueza
de especies y biomasa (Gentry
& Dodson, 1987; Hofstede et al., 1993; Krömer et al., 2005). Por otra parte, desempeñan un rol importante en el mantenimiento de la diversidad biológica y generan
microhábitats, alimento
y refugio para la fauna (Benzing, 1990).
La distribución vertical de epífitas
en estratos o zonas del árbol resulta
de las condiciones microclimáticas que ofrece el forófito y las características de las epífitas (Zotz, 2007). La zonificación más empleada es la de
Johansson (1974), que divide al forófito en cinco zonas en función de la
estructura del árbol. A medida que se asciende
por el eje vertical del forófito disminuyen la longevidad del sustrato
y, por lo tanto, la disponibilidad temporal para la colonización.
También disminuyen el grosor de las ramas, la cantidad de sustrato, la rugosidad de la
corteza y la capacidad para almacenar agua,
lo cual aumenta las diferencias en disponibilidad
hídrica impuestas por el gradiente micro-
climático (Zotz, 2007).
La distribución vertical de las epífitas dentro de un forófito está determinada por varios gradientes microambientales, donde la intensidad de la luz, la velocidad del viento y la temperatura del aire aumentan, y la humedad
del aire disminuye
desde el nivel del suelo hasta el dosel (Krömer et
al., 2007).
Los bosques húmedos tropicales en la región austral
de América incluyen
a las Yungas australes, que se extienden desde el centro-sur de Bolivia y el noroeste
de Argentina en las provincias de Jujuy,
Salta, Tucumán y el noreste de Catamarca, entre los 22° y 29° S (Cabrera
& Willink, 1980; Brown
& Kappelle, 2001).
La ecorregión de las Yungas
por sus características de humedad y precipitación
anual constituyen el ambiente
propicio para el crecimiento de plantas epífitas, especialmente de helechos (Malizia, 2003; Ceballos et al. 2016).
Diversos estudios han señalado que algunas especies
arbóreas actúan como forófitos
dominantes o preferenciales
para el establecimiento de epífitas debido
a características como,
la arquitectura del dosel, la retención de humedad y la textura de la corteza.
En los pisos altitudinales de las Yungas, se destacan como forófitos
dominantes Cedrela angustifolia Sessé &
Moc. ex DC., Tipuana tipu (Benth.) Kuntze,
Alnus acuminata Khnth, Juglans australis
Griseb., Ocotea
porphyria (Griseb.) van der Werff., Podocarpus parlatorei Pilg., entre otros (Brown,
1990; Brown et al., 2002; Malizia,
2003).
El presente trabajo tiene como objetivo
caracterizar la diversidad de epífitos vasculares
y su distribución vertical,
sobre diez forófitos
dominantes o codominantes, en las Yungas australes del noroeste argentino.
MATERIALES Y MÉTODOS
El relevamiento se realizó en cinco localidades del noroeste argentino, en un gradiente
latitudinal, comprendido entre los 23º 40’ y 24º 43’ de latitud
S. En la provincia de Jujuy: Dpto. Doctor Manuel Belgrano
Potrero de Yala (24° 6’ 48.95”,65° 28’ 13.08”), Dpto. Ledesma, Parque Nacional Calilegua
(23° 40’59.74”, 64° 54’ 5.47”),
Tiraxi (24° 0’ 18.77”, 65° 21’ 5.45”) y en la provincia
de Salta: Dpto. La Caldera,
Ruta Nacional Nº 9, ‘camino
de cornisa’, 24° 30’ 19.54”, 65° 18’ 52.71”)
y Dpto. Capital, Quebrada
de San Lorenzo (24° 43’ 11.45”,
65° 30’ 56.64”) (Fig. 1).
Se seleccionaron 10 forófitos por su dominancia y porte: Anadenanthera colubrina var. cebil
(Griseb.) Altschul. (cebil colorado), Cedrela angustifolia (cedro jujeño), Ocotea porphyria (laurel de la falda), Enterolobium contortisiliquum (Vell.) Morong.
(pacará), Erythrina falcata Benth. (ceibo jujeño), Gleditsia amorphoides (Griseb.) Taub. (espina de corona), Nectandra angusta Rohwer (laurel), Parapiptadenia excelsa
(Griseb.) Burkart (horco cebil),
Tipuana tipu (tipa), y Zanthoxylum coco Gill. (coco, cocucho).
Por cada especie de forófito se muestrearon
cinco a diez individuos, considerando ejem- plares con diámetro a la altura
del pecho (DAP)
mayor de 30 cm. Para cada árbol se completó una planilla registrando la distribución vertical de epífitos en cuatro zonas del hospedador:
tronco (Tr), copa interna tanto en ramas (CIr) como en horquetas (CIh) y copa externa
(CEx) (Fig. 2). Posteriormente, se calcularon
los porcentajes de presencia de cada especie epífita en las distintas zonas del
forófito
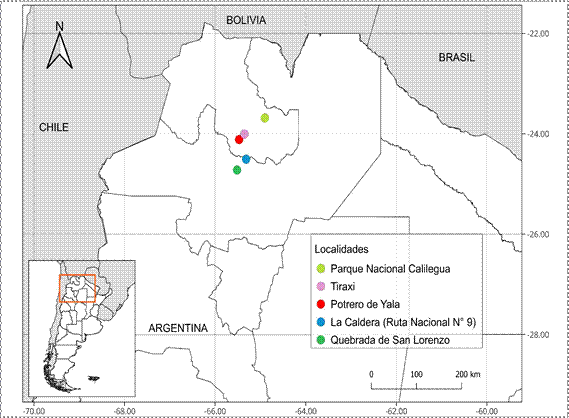
Figura 1.
Localidades del noroeste argentino donde se realizó el estudio de la diversidad epifítica.
utilizando tablas dinámicas y funciones estadísticas
básicas de Microsoft Excel.
Los ejemplares de referencia fueron depositados en el Herbario
MCNS (Facultad de Ciencias
Naturales, Universidad Nacional de Salta, Argentina). Debido
a la gran cantidad de ejemplares registrados, se citan solamente especies de helechos representativos (Tabla 1).
Para el análisis
de los datos se aplicó un
Análisis de Componentes Principales (PCA) con el objetivo de evaluar si la composición de especies de epífitos varía entre los diferentes forófitos muestreados. Para ello, se construyó
una matriz de presencia/ausencia, en la que las filas correspondieron a los individuos de forófitos
y las columnas a las especies
de epífitos registradas. La presencia de una especie en un forófito
fue codificada como “1”, y su ausencia como “0”.
El análisis se llevó a cabo en el entorno RStudio (versión 3.6), utilizando la función
|
Especies
|
Localidad
|
Colector/es
|
|
Asplenium auritum Sw.
|
Prov. Salta: Dpto. Capital, Quebrada de San Lorenzo
|
Jaimez & Martínez 14 .
|
|
Asplenium cuspidatum Lam.
|
Prov. Salta: Dpto. Capital, Quebrada de San Lorenzo
|
Jaimez & Martínez 13 .
|
|
Asplenium praemorsum
Sw.
|
Prov. Salta: Dpto. Capital, Quebrada de San Lorenzo
|
Jaimez & Martínez 15 .
|
|
Cystopteris diaphana (Bory) Blasdell
|
Prov. Jujuy: Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al. 21
|
|
Dryopteris
patula
(Sw.) Underw.
|
Prov. Jujuy: Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al. 33
|
|
Campyloneurum aglaolepis (Alston) Sota
|
Prov. Jujuy: Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al. 48
|
|
Campyloneurum angustifolium Fée
|
Prov. Jujuy: Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al. 49
|
|
Campyloneurum lorentzii (Hieron.) Ching
|
Prov. Salta: Dpto. Capital, Quebrada de San Lorenzo
|
Jaimez & Martínez 12
|
|
Microgramma squamulosa (Kaulf.) de la
Sota
|
Prov. Salta: Dpto. La Caldera, camino de cor- nisa
|
Jaimez et
al. 40
|
|
Pecluma barituensis O.G.Martínez &
de la Sota
|
Prov. Jujuy: Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al.50
|
|
Pecluma filicula (Kaulf.) M.G.Price
|
Prov. Salta Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al.52
|
|
Pecluma oranense de la Sota
|
Prov. Salta Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al.54
|
|
Phlebodium areolatum
(Willd.) J.Sm.
|
Dpto. La Caldera,
camino de cornisa,
|
Jaimez et
al.55
|
|
Pleopeltis macrocarpa Kaulf.
|
Prov. Jujuy. Dpto. Ledesma, Parque Nacional Calilegua
|
Jaimez et
al. 37
|
|
Pleopeltis minima
(Bory)
J.Prado & R.Y.Hirai
|
Prov. Salta: Dpto. La Caldera, camino de cor- nisa
|
Jaimez et
al. 80
|
|
Pleopeltis pleopeltidis (Fée) de la
Sota
|
Prov. Jujuy: Dpto. Doctor Manuel Belgrano, Parque provincial
potrero de Yala.
|
Jaimez et
al. 70
|
|
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Prov. Jujuy: Dpto. Doctor Manuel Belgrano, Tiraxi
|
Jaimez et
al. 78
|
|
Serpocaulon australe
D.Sanín, J.C.Ospina, I.O.Moura &
Salino
|
Prov. Salta: Dpto. Capital, Quebrada de San Lorenzo
|
Jaimez et
al. 100
|
Tabla 1. Ejemplares representativos del material estudiado.
prcomp() del paquete base stats, al ser datos binarios no requieren estandarización adicional. Se examinaron los primeros
componentes principales para identificar
patrones de agrupamiento entre los forófitos
según su composición epifítica, y se
visualizaron los resultados mediante un Biplot (RStudio
Team, 2025). Este análisis
permitió explorar la variación
multivariada y determinar si ciertas especies
de forófitos tienden a albergar conjuntos específicos de epífitos.
RESULTADOS
En este estudio se registró un total de 47
especies epífitas vascularesentreelloshelechos
y angiospermas (Fig. 2, Tabla 2). Los helechos
epífitos son 18 especies correspondientes a cuatro familias:
Aspleniaceae (3 especies), Cystopteridaceae (1), Dryopteridaceae (1) y
Polypodiaceae (13). La familia más diversa
de helechos fue Polypodiaceae (Fig. 2), con 9 géneros y 13 especies, y el género
mejor representado es Pleopeltis.
Las especies epífitas
presentes en la ma-
yoría de los forófitos, consideradas especies generalistas, son Campyloneurum aglaolepis (Alston) de la Sota, Microgramma squamulosa (Kaulf.) de la Sota y Pleopeltis tweedieana (Hook.) A.R.Sm., mientras
que las epífitas ocasionales son Asplenium lorentzii Hieron., Cystopteris diaphana (Bory) Blasdell y Dryopteris patula (Sw.) Underw.
(Fig. 3).
Los forófi
Cedrela angustifolia
y Ocotea porphyria
tienen la mayor diversidad de helechos
epífi
(nueve especies), mientras que Anadenanthera colubrina var. cebil y Zanthoxylum coco presentan la menor diversidad (tres especies cada uno). Se registraron 29 especies de angiospermas
epífi
de las familias Bromeliaceae, Cactaceae, Orchidaceae y Piperaceae, coexistiendo con especies de Polypodiaceae (Tabla 2, Fig. 2).
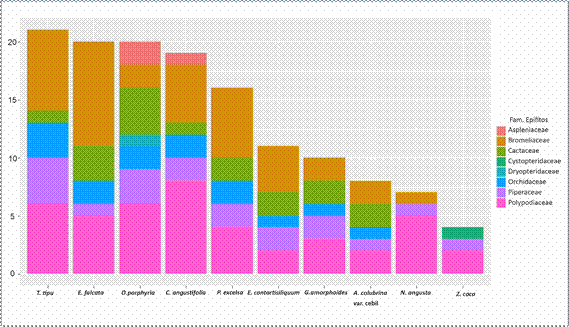
Figura 2. Gráfico de frecuencias absolutas, mostrando la diversidad de familias de epífitos presentes
sobre cada forófito.

Figura 3. Epífitos. A. helechos epífitos sobre Erythrina falcata.
B. epífitos sobre
Tipuana tipu. C. Microgramma squamulosa. D. Campyloneurum
aglaolepis. E. Pleopeltis tweedieana.
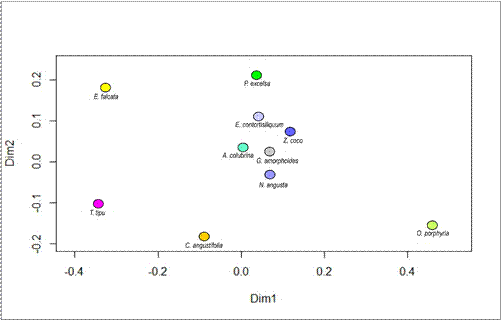
Figura 4. Biplot
de presencia/ausencia de epífitos sobre 10 forófitos.
Los forófitos con
mayor número de epífitas,
considerando helechos y angiospermas, son Tipuana tipu (23 especies),
Erythrina falcata y Ocotea porphyria (20
especies cada uno) (Figs. 2, 3A, B) y los de menor
número de epífitos, Nectandra angusta (7 especies)
y Zanthoxylum coco (4 especies) (Fig. 2).
El biplot resultante del análisis de componentes principales (Fig. 4) muestra
la variabilidad en la presencia de epífitos en los
distintos forófitos. Se observa una agru-
pación central de cinco especies
arbóreas Anadenanthera colubrina var. cebil, Entero-
lobium contortisiliquum, Nectandra angusta, Gleditsia
amorphoides y Zanthoxylum coco, que presentan patrones similares en la presencia de epífitos. Estos forófitos se caracterizan por presentar el menor número de especies epífitas (3 a 11). Por otro lado, los cinco forófitos ubicados de manera periférica
en el biplot, alejados
del grupo central,
Tipuana tipu, Erythrina falcata, Ocotea
porphyria, Cedrela angustifolia y Parapiptadenia excelsa, presentan patrones distintivos en la composición epifítica y concentran la mayor diversidad (16 a 23 especies).
El análisis de la distribución vertical de los epífitos (Tabla 3) revela que la
copa interna (CIr + CIh) alberga el 76 % de la diversidad epifítica total, con un
49 % en las ramas de la copa interna y un 27 % en las horquetas de la copa interna.
El porcentaje restante se registra principalmente en el tronco (18 %) y, en menor
medida, en la copa externa (6 %) (Fig. 5).
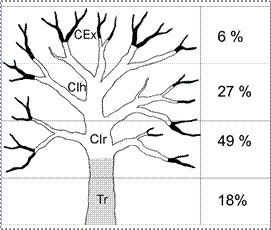
Figura 5. Esquema de forófito, donde se indican
las zonas de distribución vertical
de los epífitos: tronco (gris), copa interna
(blanco) y copa externa (negro).
Tr: tronco. CIr: copa interna, ramas. CIh: copa interna,
horquetas. CEx: copa externa.
|
Forófito
|
Helechos epífitos
|
Epífitos acompañantes
|
|
Anadenanthera
colubrina var.
cebil (Griseb.) Altschul
|
Microgramma squamulosa (Kaulf.) de la
Sota
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Orchidaceae sp.1,
Peperomia tetraphylla Hook.
& Arn.,
Rhipsalis floccosa Salm-Dyck, R. lumbricoides Lem., Tillandsia capillaris Ruiz & Pav., Vriesea friburgensis Mez
|
|
Cedrela angustifolia
Sessé &
Moc. ex Dc.
|
Asplenium cuspidatum Lam.
Campyloneurum aglaolepis (Alston) de la
Sota Campyloneurum angustifolium
Fée Campyloneurum lorentzii
(Hieron.) Ching Microgramma squamulosa (Kaulf.) de
la Sota Pecluma barituensis O.G.Martínez & de
la Sota Pecluma oranense de la Sota
Pleopeltis pleopeltidis (Fée) de la
Sota
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Aechmea distichantha
Lem., Gomesa
bifolia (Sims) M.W.Chase & N.H.Williams, Orchidaceae sp.2., Peperomia lorentzii C.DC.
ex Kuntze, P. tetraphylla Hook.
& Arn.,
Rhipsalis lumbricoides Lem.,Tillandsia australis Mez., T. recurvata (L.) L., T. capillaris Ruiz
& Pav., T. tenuifolia L.
|
|
Enterolobium
contortisiliquum (Vell.) Morong.
|
Microgramma squamulosa (Kaulf.) de la
Sota
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Gomesa bifolia
(Sims) M.W.Chase & N.H.Williams,
Peperomia lorentzii C.DC. ex Kuntze, P. tetraphylla Hook.
& Arn.,
Rhipsalis monacantha Griseb.
R. lumbricoides Lem.,
Tillandsia minutiflora
Donadio, T. didisticha
Baker
T. tenuifolia L.,
Vriesea friburgensis Mez
|
|
Erythrina falcata
Benth.
|
Campyloneurum aglaolepis
(Alston) de la
Sota Microgramma squamulosa (Kaulf.) de la
Sota Pleopeltis minima
(Bory) J.Prado & R.Y.Hirai Pleopeltis tweedieana (Hook.) A.R.Sm.
Serpocaulon australe D.Sanín,
J.C.Ospina, I.O.Moura &
Salino
|
Aechmea distichantha
Lem., Gomesa
bifolia (Sims) M.W.Chase & N.H.Williams, Orchidaceae sp.2, Peperomia lorentzii C.DC. ex Kuntze, Pheiffera ianthothele F.A.C.Weber, Rhipsalis baccifera
(Sol.) Stearn, R.
lumbricoides Lem., Tillandsia
bandensis Baker, T. minutiflora Donadio,
T. capillaris Ruiz
& Pav.,
T. diaguitensis A.Cast., T. schreiteri Lillo &
A.Cast.,
T. sphaerocephala Baker, T. tenuifolia
L., Vriesea friburgensis Mez
|
|
Gleditsia amorphoides (Griseb.) Taub.
|
Campyloneurum lorentzii
(Hieron.) Ching
Microgramma squamulosa (Kaulf.)
de la Sota Pleopelits tweedieana (Hook.) A.R.Sm.
|
Gomesa bifolia
(Sims) M.W.Chase & N.H.Williams,
Peperomia lorentzii C.DC. ex Kuntze, P. tetraphylla Hook.
& Arn.,
Rhipsalis lumbricoides Lem.
R. monacantha Griseb., Tillandsia capillaris Ruiz
& Pav.,
Vriesea friburgensis Mez
|
|
Nectandra angusta
Rohwer
|
Campyloneurum aglaolepis
(Alston) Sota
Microgramma squamulosa (Kaulf.)
de la Sota Pecluma filicula (Kaulf.) M.G.Price
Pleopeltis minima
(Bory)
J.Prado & R.Y.Hirai
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Peperomia lorentzii C.DC. ex Kuntze,
Tillandsia recurvata
(L.) L.
|
|
Ocotea porphyria (Griseb.) van der Werff.
|
Asplenium auritum Sw.
Asplenium praemorsum Sw.. Campyloneurum aglaolepis
(Alston) Sota
Campyloneurum angustifolium Fée Campyloneurum
lorentzii (Hieron.) Ching
Dryopteris patula (Sw.) Underw.
Microgramma squamulosa (Kaulf.) de la
Sota
Pleopeltis macrocarpa Kaulf.
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Gomesa bifolia
(Sims) M.W.Chase & N.H.Williams, Orchidaceae sp. 2, Peperomia collinsii Villa, Peperomia inaequalilimba
C.DC., Peperomia lorentzii C.DC. ex Kuntze,
Pfeiffera ianthothele F.A.C.Weber,
Rhipsalis monacantha Griseb., Rhipsalis lorentziana Griseb., Rhipsalis lumbricoides
Lem., Tillandsia didisticha
Baker Tillandsia recurvata (L.) L.
|
|
Parapiptadenia
excelsa (Griseb.) Burkart
|
Campyloneurum aglaolepis
(Alston) Sota
Microgramma squamulosa (Kaulf.)
de la Sota Pleopeltis pleopeltidis (Fée) de la
Sota Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Aechmea distichantha
Lem., Gomesa
bifolia (Sims) M.W.Chase & N.H.Williams, Orchidaceae sp. 1, Peperomia lorentzii C.DC.
ex Kuntze, P. tetraphylla Hook.
& Arn.,
Rhipsalis floccosa Salm-Dyck, R.
monacantha
Griseb., Tillandsia
minutiflora Donadio, T. capillaris Ruiz
& Pav., T.
didisticha Baker,
T. sphaerocephala Baker, T. usneoides
(L.) L.
|
|
Tipuana tipu (Benth.) Kuntze
|
Campyloneurum aglaolepis
(Alston) Sota
Microgramma squamulosa (Kaulf.)
de la Sota Phlebodium areolatum
(Willd.) J.Sm.
Pleopeltis minima
(Bory)
J.Prado & R.Y.Hirai Pleopeltis pleopeltidis (Fée) de
la Sota Pleopeltis tweedieana (Hook.) A.R.Sm.
|
Aechmea distichantha
Lem., Gomesa
bifolia (Sims) M.W.Chase & N.H.Williams, G. viperina (Lindl.) M.W.Chase & N.H.Williams, Orchidaceae sp.1, Peperomia lorentzii C.DC.
ex Kuntze, P. tetraphylla Hook.
& Arn., P. theodori Trel., Pheiffera ianthothele F.A.C.Weber, Rhipsalis floccosa Salm-Dyck, R. lumbricoides Lem., Tillandsia
australis Mez, T.
bandensis Baker, T. capillaris Ruiz & Pav., T. didisticha
Baker, T. recurvata (L.) L., T. tenuifolia L., Vriesea friburgensis Mez
|
|
Zanthoxylum coco Gill.
|
Campyloneurum aglaolepis
(Alston) Sota
Pleopeltis tweedieana (Hook.) A.R.Sm. Cystopteris
diaphana (Bory) Blasdell
|
Peperomia tetraphylla Hook. & Arn.
|
Tabla 2.
Diversidad epifítica total de helechos y epifitas acompañantes
por especie de forófito.
|
Familias
|
Especies de helechos
epífitos
|
Distribución vertical de epífitos
|
|
Tr
|
CIr
|
CIh
|
CEx
|
|
Aspleniaceae
|
Asplenium auritum Sw.
|
x
|
x
|
|
|
|
Aspleniaceae
|
Asplenium cuspidatum Lam.
|
|
x
|
|
|
|
Aspleniaceae
|
Asplenium praemorsum
Sw.
|
x
|
x
|
|
|
|
Cystopteridaceae
|
Cystopteris diaphana (Bory)
Blasdell
|
|
x
|
|
|
|
Dryopteridaceae
|
Dryopteris patula (Sw.) Underw.
|
|
|
x
|
|
|
Polypodiaceae
|
Campyloneurum aglaolepis (Alston) Sota
|
x
|
x
|
|
|
|
Polypodiaceae
|
Campyloneurum angustifolium Fée
|
|
x
|
x
|
|
|
Polypodiaceae
|
Campyloneurum lorentzii (Hieron.) Ching
|
|
x
|
|
|
|
Polypodiaceae
|
Microgramma squamulosa (Kaulf.)
de la Sota
|
x
|
x
|
x
|
x
|
|
Polypodiaceae
|
Pecluma barituensis O.G.Martínez &
de la Sota
|
|
x
|
x
|
|
|
Polypodiaceae
|
Pecluma filicula (Kaulf.)
M.G.Price
|
x
|
|
|
|
|
Polypodiaceae
|
Pecluma oranense de la Sota
|
|
x
|
x
|
|
|
Polypodiaceae
|
Phlebodium areolatum
(Willd.) J.Sm.
|
|
x
|
x
|
|
|
Polypodiaceae
|
Pleopeltis macrocarpa Kaulf.
|
|
x
|
x
|
x
|
|
Polypodiaceae
|
Pleopeltis minima
(Bory) J.Prado & R.Y.Hirai
|
x
|
x
|
|
|
|
Polypodiaceae
|
Pleopeltis pleopeltidis (Fée)
de la Sota
|
|
x
|
x
|
|
|
Polypodiaceae
|
Pleopeltis tweedieana (Hook.) A.R.Sm.
|
|
x
|
x
|
|
|
Polypodiaceae
|
Serpocaulon australe
D.Sanín, J.C.Ospina, I.O.Moura & Salino
|
|
x
|
|
|
Tabla 3. Diversidad y distribución vertical de los helechos epífitos sobre diez forófitos; Tr: tronco; CIr: copa interna,
ramas; CIh: copa interna, horquetas. CEx: copa externa.
DISCUSIÓN Y CONCLUSIÓN
El epifitismo de los bosques nublados de la ecorregión de las Yungas,
presenta un pico de diversidad entre los 1000 y 2000 m
s.n.m. (Gentry & Dodson,
1987; Brown, 1990), así Krömer et al. (2005) y Ceballos (2023) mencionan
que en las Yungas
la mayor diversidad de epífitas se encuentra a los
1500 m s.n.m.
Nuestros resultados muestran
que los helechos epífitos presentan la mayor
diversidad de especies en la región de las Yungas,
lo que resalta su importancia dentro
de la comunidad epifítica local. En este sentido,
de la Sota (1971) destaca que las pteridofitas epífitas desempeñan un papel clave
en los ecosistemas tropicales, no solo por su contribución a la
diversidad florística, sino también por su
función en el ciclo hídrico
y la dinámica de
nutrientes.
Otros estudios realizados en el noroeste argentino registraron una diversidad de 47 especiesepífitas,
principalmentedepteridófitas y bromeliáceas, en el Parque
Nacional El Rey, provincia de Salta (Brown, 1990). Asimismo,
Malizia (2003)
reportó 23 especies de epífitos y plantas
trepadoras en la Reserva El Nogalar, en Jujuy, donde Polypodiaceae resultó
la
familia más diversa. De manera coincidente con nuestros resultados, las familias Polypodiaceae y Bromeliaceae fueron las que presentaron la mayor diversidad
de especies en ambas áreas de las Yungas.
Las especies epífitas
de Polypodiaceae más frecuentes en los forófitos analizados son Campyloneurum aglaolepis, Microgramma squamulosa y Pleopeltis tweedieana. Estos resultados concuerdan con los hallazgos
de otros estudios realizados en áreas de las
Yungas, como los de Ceballos et
al. (2016) en el Parque Sierra de San Javier de la provincia
de Tucumán, y Brown (1990)
en el Parque Nacional El
Rey de la provincia de Salta.
La elevada
diversidad de helechos epífitos
y su éxito en la colonización de distintos sustratos pueden explicarse, en gran medida, por sus adaptaciones morfológicas y fisiológicas.
Entre las características más relevantes se encuentran los rizomas largamente rastreros, escamas, pelos y ceras que contribuyen a la retención de humedad y adhesión al sustrato. Además,
este grupo presenta mecanismos como el enrollamiento y la curvatura
de la lámina foliar, e incluso
la pérdida total de las frondes durante
períodos de sequía, estrategias que les permiten
tolerar condiciones ambientales extremas
(de la Sota, 1971; Tryon, 1986).
Otra adaptación relevante de los helechos epífitos
es la producción de esporas
con alta capacidad de dispersión y viabilidad. Las esporas pueden germinar en dos o tres días tras su liberación, o bien permanecer viables durante meses
e incluso años en el sustrato, lo que
favorece su establecimiento en una amplia
gama de condiciones ambientales; estas
estrategias contribuyen significativamente al
éxito de los helechos epífitos
en las comunidades de dosel, en comparación con otros grupos de epífitos (Tryon, 1986; Raghavan, 1989; Barrington, 1993).
La vegetación epifítica vascular
es alta- mente dependiente de la permanencia del bosque, la que puede
estar limitada por la corteza agrietada y follaje caducifolio de los forófitos
(Martín et al.,
2008). Además, Zotz & Andrade (2002) mencionan
que la riqueza y abundancia de epífitas en distintos ambientes están determinadas en parte por factores ecológicos (disponibilidad de agua y luz) y condiciones micro y macroclimáticas.
Cortés-Anzúres et
al. (2020), mencionan que son pocas las epífitas que pueden colonizar a la mayoría de los forófitos. Algunas como las orquídeas
aparecen en pocos forófitos y/o tienen baja densidad poblacional, lo que sugiere
que están fuertemente limitados por los forófitos. Sugden & Robins (1979) afirman
que las epífitas son más abundantes en aquellos forófitos muy
ramificados hacia todos los ángulos, con ramas horizontales y grandes copas, tal como se observa en Tipuana tipu “tipa” y Ocotea porphyria “laurel de la falda”.
Otro estudio sobre epifitismo en regiones
neotropicales destaca que ciertas especies arbóreas albergan una mayor riqueza
de epífitos, debido
a características como la
rugosidad de la corteza, la arquitectura del dosel y la capacidad
de retención de humedad
(de la Sota, 1971). De manera concordante, en nuestro trabajo los forófitos con alta diversidad de epífitos son Cedrela angustifolia, Erythrina falcata, Ocotea porphyria y Tipuana tipu, árboles
con corteza rugosa,
de ramificación densa y copas amplias.
Estas características
estructurales resultan fundamentales para el
establecimiento y la persistencia de comunidades epifíticas al ofrecer una mayor
superficie disponible, retención
de agua y microhábitats favorables (de la Sota, 1971).
Los registros de diversidad de epífitos asociados a Ocotea porphyria varían según la localidad estudiada. Brown (1990) registró 14 especies epífitas sobre esta
especie en el Parque Nacional
El Rey, mientras que Ceballos et al. (2016)
reportaron 16 especies en un bosque
subtropical del Parque
Sierra de San Javier,
en la provincia de Tucumán. En el presente
estudio se encontraron 20 especies epífitas sobre Ocotea porphyria, lo que representa
la mayor cantidad
de epífitos para esta especie en la zona de estudio.
La distribución vertical
de los epífitos, muestra que el mejor soporte para su
establecimiento es la copa interna, probablemente debido a la acumulación de materia orgánica sobre la corteza
como ocurre en otras leñosas (Benzing, 1990; Barthlott et al.,
2001; Hernández-Rosas, 2001; Hernández-
Rosas, 2004), además de la humedad que se
genera la copa interna de los árboles, forman
un microclima ideal para estas plantas (de la
Sota, 1971).
Los resultados obtenidos en este trabajo
no permiten establecer preferencias específicas de las epífitas
por un determinado tipo de corteza,
posición topográfica o exposición de los forófitos, resultados también registrados
por otros autores (Hietz & Briones, 1998; Barthlott et al., 2001;
Martín et al.,
2008).
Se concluye que la diversidad de epífitos sobre diez forófitos de Yungas en el noroeste
argentino es relativamente alta (47 especies),
con 18 especies de helechos, en relación a otros
estudios comprendidos en un rango altitudinal semejante, entre los 800-1800 m s.n.m.,
zona reconocida por la alta diversidad
de epífitos (Malizia, 2003; Ceballos
et al. 2016; Ceballos, 2023).
Es fundamental realizar investigaciones en profundidad sobre las epífi de las Yungas australes de Sudamérica para identifi con precisión los factores que infl en su propagación, establecimiento y desarrollo, para aportar
datos ecofi de interés para promover
estrategias efectivas de conservación.
AGRADECIMIENTOS
Agradecemos a Janet Chambi
y Marcelo Gerónimo por la colaboración y asistencia técnica
durante el desarrollo de este trabajo. A Olga Martínez por la lectura
crítica del manuscrito y a los revisores anónimos por las observaciones y recomendaciones realizadas. Este trabajo
fue subsidiado por el Consejo
de Investigación de la Universidad Nacional
de Salta, Argentina.
REFERENCIAS
Barrington, D. S. (1993).
Ecological and histo- rical factors in fern biogeography. Journal of Biogeography, 20, 275-280. https://doi. org/10.2307/2845635
Barthlott, W., Schmith-Neuerburg V., Nieder J., & Engwald,
S. (2001). Diversity and abundance of vascular epiphytes:
a comparision of a
secondary vegetation and primary montane rainforest in the Venezuelan Andes. Plant Ecology, 152, 145-156.
https://doi. org/10.1023/A:1011483901452
Benzing, D. H. (1990).
Vascular Epiphytes
Cambridge University Press. Cambridge.
Journal of tropical ecology, 12, 354.
Bonnet, A., Curcio, G. R., Lavoranti, O. J.,
& Galvão, F. (2011). Flora epifítica
vascular em três unidades vegetacionais do Rio Tibagi, Paraná,
Brasil. Rodriguésia, 62(3), 491-498.
https://doi.org/10.1590/2175-7860201162305
Brown, A. D. (1990).
El epifitismo en las selvas
montanas del Parque Nacional “El Rey”,
Argentina: Composición florística y patrón de distribución. Revista de Biología
Tropical, 38 (2A),
155-166. https://archivo.revistas.ucr. ac.cr/index.php/rbt/article/view/25037
Brown, A. D. & Kappelle, M. (2001). Introducción a los bosques nublados del neotrópico:
una síntesis regional. En M. Kappelle, & D. Brown (Eds.), Bosques
nublados del neotrópico (pp. 25-40). Instituto Nacional de Biodiversidad (INBio), IMBIO, Santo Domingo de Heredia, Costa Rica.
Brown, A. D., Grau, A., Lomáscolo, T., & Gasparri, N. I. (2002).
Una estrategia de conservación para las selvas subtropicales de montaña
(Yungas) de Argentina. Ecotropicos, 15(2), 147-159.
Cabrera, A., & Willink, A. (1980). Biogeografía de América Latina. Secretaría General
de la Organización de los Estados
Americanos. Programa Regional de
Desarrollo Científico y Tecnológico. Serie de Biología.
Monografía, (13).
Ceballos, S. J., Chacoff,
N. P., & Malizia,
A. (2016). Interaction network of vascular epiphytes and trees in a subtropical forest. Acta Oecologica, 77, 152-159.
https://doi. org/10.1016/j.actao.2016.10.007
Ceballos, S. J. (2023).
Vascular epiphytes in Argentinian Yungas: distribution, diversity, and ecology.
The Botanical Review, 89, 91-113.
https://doi.org/10.1007/s12229-
022-09281-7
Cortés-Anzúres, B. O., Corona-López, A. M., Damon, A., Mata-Rosas, M., & Flores- Palacios, A. (2020). Phorophyte type determines epiphyte-phorophyte network structure in a Mexican oak forest. Flora, 272, 151704. https://doi.org/10.1016/j. flora.2020.151704
de la Sota, E. R. (1971). El epifitismo
y las pteridofitas en Costa Rica (América Central). Nova Hedwigia, 21, 401-465.
Gentry, A. H., & Dodson,
C. H. (1987). Diversity and biogeography of Neotropical
vascular epiphytes. Annals of the Missouri Botanical Garden, 74, 205-233.
https://doi. org/10.2307/2399395
Hernández-Rosas, J. (2001).
Ocupación de los portadores por epífitas vasculares en un bosque húmedo tropical del Alto Orinoco, Edo. Amazonas, Venezuela. Acta Científica Venezolana, 52, 292-303.
Hernández-Rosas, J. (2004). Características del substrato
de plantas del dosel de un
bosque húmedo tropical de tierras
bajas (Alto Orinoco, Venezuela). Acta Científica Venezolana, 55, 35-43.
Hietz, P., & Briones,
O. (1998). Correlation between water relations
and within-canopy distribution of epiphytic ferns in a Mexican
cloud forest. Oecologia, 114, 305-316. https:// doi.org/10.1007/s004420050452
Hirata, A., Kamijo,
T., & Saito, S. (2008). Host trait preferences and distribution of vascular epiphytes. In Van der. Valk (Ed.), Forest Ecology.
Springer, Dordrecht.
Hofstede, R. G., Wolf, J. H., & Benzing,
D. H. (1993). Epiphytic
biomass and nutrient
status of a Colombian upper montane rain forest.
Selbyana, 14, 37-45.
Ibisch, P. L., Boegner A., Nieder, J., & Barthlott, W. (1996). How diverse are Neotropical epiphytes? An analysis based on the
“Catalogue of the Flowering Plants and Gymnosperms of Peru”. Ecotropica, 2, 13-28.
Johansson, D. (1974).
Ecology of vascular epiphytes in West African rain forest. Acta
Phytogeographica Suecica, 59, 1-136.
Kersten, R. A. (2010). Epífitas
vasculares– Histórico, participação taxonômica e aspectos relevantes, com ênfase na Mata Atlântica. Hoehnea, 37(1),
9-38. https://doi.org/10.1590/ S2236-89062010000100001
Krömer, T., Kessler, M., Gradstein, S. R., &
Acebey,
A. (2005). Diversity
patterns of vascular epiphytes along an elevational gradient in the Andes. Journal of Biogeography, 32, 1799-1809. https://doi.org/10.1111/j.1365- 2699.2005.01318.x
Krömer, T., Kessler, M., & Gradstein, S. R. (2007).
Vertical stratification of vascular epiphytes in submontane and montane forest of
the Bolivian Andes: the importance
of the understory. Plant Ecology, 189, 261-278. https://doi.org/10.1007/s11258-006-9182-8
Laube, S., &
Zotz, G. (2006).
Neither host-specific nor random:
vascular epiphytes
on three tree species in a Panamanian
lowland forest. Annals of Botany, 97(6), 1103-1114. https:// doi.org/10.1093/aob/mcl067
Malizia, A. (2003).
Host tree preference of vascular
epiphytes and climbers in a subtropical
montane cloud forest of northwest
Argentina. Selbyana, 24, 196-205.
https://www.jstor.org/ stable/41760133
Martín, J. S., Espinosa, A., Zanetti, S., Hauenstein,
E., Ojeda, N., & Arriagada, C. (2008). Composición y estructura de la vegetación epífita vascular en un bosque primario
de Olivillo (Aextoxicon punctatum
R. et P.) en el sur de Chile.
Ecología Austral, 18,1-11.
Raghavan, V. (1989). Developmental Biology
of Ferns. Pp.331.
Cambridge Press.
RStudio Team. (2025). RStudio:
Integrated Development for R. RStudio,
PBC, Boston, MA.
Sugden, A. M., & Robins,
R. J. (1979). Aspects of the ecology of vascular epiphytes in Colombian could forests,
I. The distribution of the epiphytic
flora. Biotropica 11(3), 173- 188. https://doi.org/10.2307/2388037
Tryon, R. M. (1986). The biogeography of species, with special reference
to ferns. Botanical Review, 52, 117-156.
https://doi.org/10.1007/ BF02860999
Wagner, K., Mendieta-Leiva, G., & Zotz, G. (2015). Host specificity in vascular epiphytes: a review of methodology, empirical evidence
and potential mechanisms. AoB plants, 7, plu092. https://doi.org/10.1093/ aobpla/plu092
Wolf, J. H. D. (1993)
Epiphyte communities of tropical montane
rain forests in the northern Andes. I. Lower montane
communities. Phytocoenologia 22, 1–52.
Zotz, G. & Andrade, J. L. (2002).
La ecología y fisiología de las epífitas
y las hemiepífitas. En M. R. Guariguata, & G. Kattan (Eds.),
Ecología y Conservación De Bosques Neotropicales (p. 271). Ediciones
LUR.
Zotz, G. (2007).
Johansson revisited: the spatial
structure of epiphyte assemblages. Journal of Vegetation Science, 18(1), 123-130.
https:// doi.org/10.1111/j.1654-1103.2007.tb02522.x
Zotz, G. (2013).
The systematic distribution of vascular epiphytes–a critical update. Botanical
Journal of the Linnean Society, 171(3), 453- 481. https://doi.org/10.1111/boj.12010
Zotz,
G. (2016). Plants
on plants - The
biology of vascular epiphytes. Springer, Suiza.